
12 results
Brain Stress Test for Assessing Risk for Hemodynamic Stroke
-
- Journal:
- Canadian Journal of Neurological Sciences / Volume 51 / Issue 1 / January 2024
- Published online by Cambridge University Press:
- 10 January 2023, pp. 57-63
-
- Article
- Export citation
Slowed Temporal and Parietal Cerebrovascular Response in Patients with Alzheimer’s Disease
-
- Journal:
- Canadian Journal of Neurological Sciences / Volume 47 / Issue 3 / May 2020
- Published online by Cambridge University Press:
- 13 February 2020, pp. 366-373
-
- Article
-
- You have access
- HTML
- Export citation
Radar absorption, basal reflection, thickness and polarization measurements from the Ross Ice Shelf, Antarctica
-
- Journal:
- Journal of Glaciology / Volume 61 / Issue 227 / 2015
- Published online by Cambridge University Press:
- 10 July 2017, pp. 438-446
-
- Article
-
- You have access
- HTML
- Export citation
-
Radio-glaciological parameters from the Moore’s Bay region of the Ross Ice Shelf, Antarctica, have been measured. The thickness of the ice shelf in Moore’s Bay was measured from reflection times of radio-frequency pulses propagating vertically through the shelf and reflecting from the ocean, and is found to be 576 ± 8 m. Introducing a baseline of 543 ± 7m between radio transmitter and receiver allowed the computation of the basal reflection coefficient, R, separately from englacial loss. The depth-averaged attenuation length of the ice column, 〈L〉 is shown to depend linearly on frequency. The best fit (95% confidence level) is 〈L(ν)〉= (460±20) − (180±40)ν m (20 dB km−1), for the frequencies ν = [0.100–0.850] GHz, assuming no reflection loss. The mean electric-field reflection coefficient is
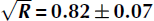 (1.7 dB reflection loss) across [0.100–0.850] GHz, and is used to correct the attenuation length. Finally, the reflected power rotated into the orthogonal antenna polarization is <5% below 0.400 GHz, compatible with air propagation. The results imply that Moore’s Bay serves as an appropriate medium for the ARIANNA high-energy neutrino detector.
(1.7 dB reflection loss) across [0.100–0.850] GHz, and is used to correct the attenuation length. Finally, the reflected power rotated into the orthogonal antenna polarization is <5% below 0.400 GHz, compatible with air propagation. The results imply that Moore’s Bay serves as an appropriate medium for the ARIANNA high-energy neutrino detector.
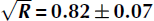 (1.7 dB reflection loss) across [0.100–0.850] GHz, and is used to correct the attenuation length. Finally, the reflected power rotated into the orthogonal antenna polarization is <5% below 0.400 GHz, compatible with air propagation. The results imply that Moore’s Bay serves as an appropriate medium for the ARIANNA high-energy neutrino detector.
(1.7 dB reflection loss) across [0.100–0.850] GHz, and is used to correct the attenuation length. Finally, the reflected power rotated into the orthogonal antenna polarization is <5% below 0.400 GHz, compatible with air propagation. The results imply that Moore’s Bay serves as an appropriate medium for the ARIANNA high-energy neutrino detector.Copyright page
-
- Book:
- The Toxicology of Carbon Nanotubes
- Published online:
- 05 July 2012
- Print publication:
- 21 June 2012, pp iv-iv
-
- Chapter
- Export citation
Contents
-
- Book:
- The Toxicology of Carbon Nanotubes
- Published online:
- 05 July 2012
- Print publication:
- 21 June 2012, pp v-ix
-
- Chapter
- Export citation
The Toxicology of Carbon Nanotubes - Half title page
-
- Book:
- The Toxicology of Carbon Nanotubes
- Published online:
- 05 July 2012
- Print publication:
- 21 June 2012, pp i-ii
-
- Chapter
- Export citation
Contributors
-
-
- Book:
- The Toxicology of Carbon Nanotubes
- Published online:
- 05 July 2012
- Print publication:
- 21 June 2012, pp x-xiv
-
- Chapter
- Export citation
The Toxicology of Carbon Nanotubes - Title page
-
-
- Book:
- The Toxicology of Carbon Nanotubes
- Published online:
- 05 July 2012
- Print publication:
- 21 June 2012, pp iii-iii
-
- Chapter
- Export citation

The Toxicology of Carbon Nanotubes
-
- Published online:
- 05 July 2012
- Print publication:
- 21 June 2012
Index
-
- Book:
- The Toxicology of Carbon Nanotubes
- Published online:
- 05 July 2012
- Print publication:
- 21 June 2012, pp 240-242
-
- Chapter
- Export citation
The contribution of chemoreflex drives to resting breathing in man
-
- Journal:
- Experimental Physiology / Volume 86 / Issue 1 / January 2001
- Published online by Cambridge University Press:
- 31 July 2001, pp. 109-116
- Print publication:
- January 2001
-
- Article
- Export citation
Changes in chemoreflex characteristics following acute carbonic anhydrase inhibition in humans at rest
-
- Journal:
- Experimental Physiology / Volume 85 / Issue 6 / November 2000
- Published online by Cambridge University Press:
- 10 January 2001, pp. 847-856
- Print publication:
- November 2000
-
- Article
- Export citation
